6.7 Séquençage des protéines
1953 fut l'année où les travaux de Crick, Watson et Rosalind Franklin nous permirent de visualiser la molécule d'ADN sous la forme hélicale qui nous est maintentant familière. Cette date devrait aussi être remémorée comme celle où où Frederick Sanger décrivit la séquence des résidus dans une molécule d'insuline.
Le séquençage protéique est un procédé classiquement décrit en six étapes.
1- Détermination de la composition en acides aminés.
2- Identification des bouts.
3- Fragmentation de la chaîne polypeptidique.
4- Séquençage des fragments.
5- Seconde coupure de la chaîne polypeptidique avec un nouvel agent.
6- Agencement des fragments obtenus et séquencés.
1- Détermination de la composition en acides aminés. Cette étape a pour but d'évaluer la quantité de chaque acide aminé présent dans un peptide ou une protéine. On y parvient en brisant chaque lien peptidique de la protéine et en analysant ensuite le mélange obtenu par séparation chromatographique sur HPLC. Bien sûr, une fois qu'on a une idée de la nature et de la quantité des acides aminés contenus dans la protéine, il reste à en déterminer l'ordre.
2- Identification des bouts. Il existe des réactifs comme le 1-fluoro-2,4-dinitrobenzene
ou le chlorure de dansyl qui réagissent avec la partie N-terminale des protéines et leur greffent un groupement supplémentaire. Alors que les liens peptidiques sont coupés par un traitement à l'acide, ces groupements supplémentaires restent greffés à au résidu situé le plus en 5' de la protéine; on peut alors identifier quel est l'acide aminé qui porte un appendice surnuméraire.
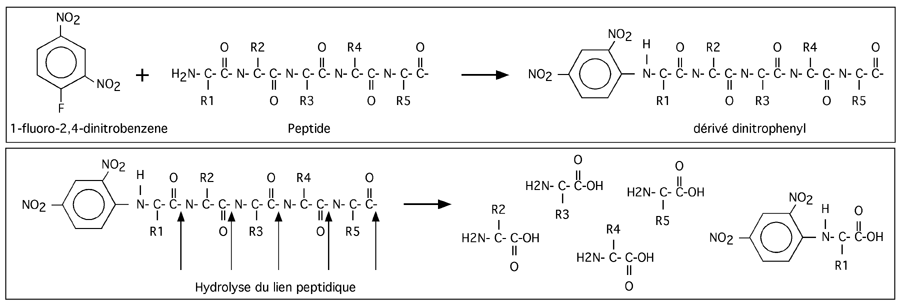
Pour l'extrémité C-terminale, on a recours à une protéolyse contrôlée avec la carboxypeptidase Y, C ou P (elles coupent le résidu situé le plus en 3 prime). Il fait alors faire attention à ne pas digérer plus d'un acide aminé à l'extrémité C-terminale.
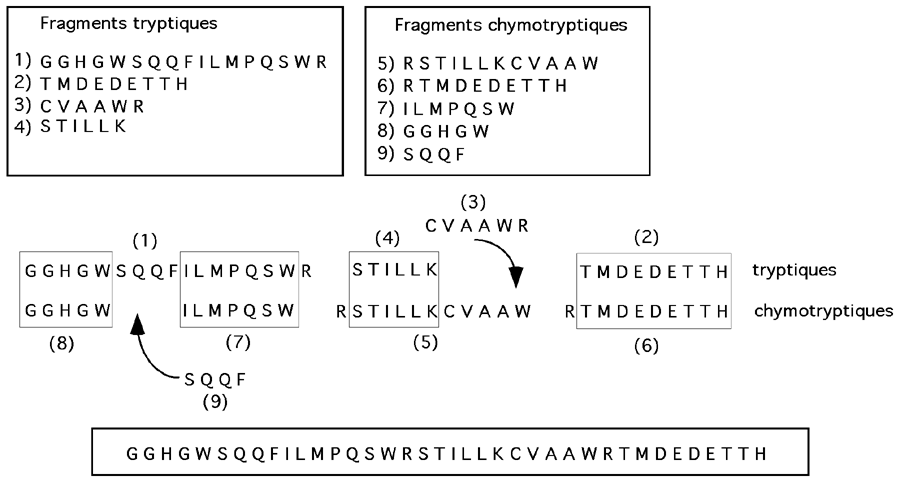
3- Fragmentation de la chaîne polypeptidique. Un traitement avec des agents chimiques ou des enzymes capables de couper la chaîne polypeptidique à des sites précis (la trypsine, par exemple, coupe après une lysine ou une arginine; la chymotrypsine après un résidu aromatique) permettra de réduire une grosse protéine en plus petits peptides. Ceux-ci pourront être séquencés séparément après séparation chromatographique.
4- Séquençage des fragments. La dégradation d'Edman permet d'enlever un par un les résidus en commençant par l'extrémité N-terminale. On arrive ainsi à déterminer la séquence de fragments d'environ 20 résidus.

5- Seconde coupure de la chaîne polypeptidique avec un nouvel agent
. Nous avons, en (3), coupé la chaîne avec un réactif ou un enzyme afin de pouvoir séquencer les fragments obtenus. Pour pouvoir enligner ces fragments, il nous faut maintenant recommencer (3) et (4) avec un nouvel échantillon de la protéine intacte et un agent différent. (Par exemple, si on a utilisé la trypsine en (3), on pourra maintenant utiliser la chymotrypsine).